Otolith contribution
to the human torsional vestibulo-ocular reflex
Introduction
The vestibular
system contributes to the stabilization of the retinal image during head movements
by means of the vestibulo-ocular reflex (VOR). Whereas
the semicircular canals are clearly involved in the control of the angular VOR
during head rotation, the role of the otolith organs is more difficult to interpret.
The otoliths respond to linear accelerations, which may result from head translation,
but also from head tilt relative to gravity. Accordingly, "translational"
and "tilt" otolith-ocular responses (OOR) have been identified, each
with different spatiotemporal properties. It is illustrative for the ambiguity
of linear acceleration and tilt as stimulus for the otoliths, that tilt responses
can be elicited by linear head motion, and translational responses can be elicited
by head rotation relative to gravity. For instance, Paige and Tomko (1991a,b)
distinguished translational OORs and tilt OORs in squirrel monkeys during horizontal
oscillation along various body axes on a linear track. Translational responses
included horizontal and vertical eye movements during motion along the interaural
axis and the dorso-ventral axis, respectively. These eye movements were most sensitive
to high-frequency oscillations and were functionally compensatory to the linear
motion of the stimulus. Tilt responses included torsional and vertical eye movements
during motion along the interaural axis and the naso-occipital axis, respectively.
In contrast to the translational responses, the tilt responses were most sensitive
to lower stimulus frequencies and were not functionally compensatory to the motion
itself. This category of eye movements was designated "tilt" OORs because
they are normally associated with inclination of the head. In man, a well-studied
tilt OOR is ocular torsion in response to static head tilt in the roll plane (Miller
1962; Belcher 1964; Bucher et al. 1992; Collewijn et al. 1985; Cheung et al. 1992).
The low-frequency dynamics of this response have been established in more detail
using linear acceleration along the interaural axis (Woellner and Graybiel 1959;
Colenbrander 1963; Hannen et al. 1966; Lichtenberg et al. 1982; De Graaf et al.
1996).
Adapted
from: Groen E, Bos JE, De Graaf B. Contribution of the otoliths to the human
torsional vestibulo-ocular reflex. Submitted to Journal of Vestibular Research
It has been assumed that ocular
torsion is determined exclusively by the shear force along the maculae of the
utricles, and not by the "swinging" of the resultant gravito-inertial
force relative to the head. Recently, however, it was suggested that the ocular
torsion response to interaural oscillation in man can be divided into a component
responding to linear motion and a component responding to the swinging resultant
gravito-inertial force (Merfeld et al. 1996). Correspondingly, the steady
state nystagmus observed during constant velocity rotation about an off-vertical
axis has been recognized as a third type of OOR, which reflects the ability
of the otoliths to estimate angular velocity from the rotation of the gravito-inertial
vector relative to the head (Guedry 1965; Benson and Bodin 1966; Angelaki and
Hess 1996a).
Basically, linear motion and
constant velocity rotation about an off-vertical axis are unnatural conditions
for tilt OORs such as ocular torsion. The findings obtained with these methods
suggest that otolith-induced ocular torsion does exist during rotations of the
head in the roll plane, when the semicircular canals are activated simultaneously.
Nevertheless, there seems to be no decisive evidence that such is the case in
humans. Collewijn et al. (1985) studied ocular torsion during active
head roll, but did not determine the contribution of the otoliths. Vieville
and Masse (1987) made an attempt to isolate the contribution of the otoliths
from that of the semicircular canals, by measuring ocular torsion during voluntary
head movements in the roll plane (in only one subject), once with the head erect,
and once with the head prone. In the head-erect condition the motion was about
an earth-horizontal axis, modulating inputs from the otoliths and the semicircular
canal. In the head-prone condition the motion was orthogonal to gravity, only
stimulating the semicircular canals. In a frequency range from 0.1 to 0.4 Hz,
the response gain was slightly higher and the phase lead smaller when the head
was erect compared to when the head was prone. Conversely, Tweed et al.
(1994) did not find a difference in three-dimensional VOR dynamics for oscillations
of 0.3 Hz about an earth-horizontal axis and about an earth-vertical axis. However,
taking the low-frequency characteristics of tilt OORs into account, a frequency
of 0.3 Hz may have been too high to reveal an otolith effect.
The goal of the present study
was to investigate whether the otoliths improve the low-frequency dynamics of
the torsional VOR in human subjects. We focused on the torsional VOR because
it is considerably weaker than the horizontal and vertical VOR (Crawford and
Vilis 1991; Tweed et al. 1994), suggesting that especially ocular torsion
could benefit from assistance of the otoliths. We compared the response to passive
sinusoidal body roll about an earth-horizontal axis and an about earth-vertical
axis (in upright and supine body orientation, respectively) in the frequency
range from 0.05 to 0.4 Hz. At very low frequencies the canal-induced VOR normally
becomes less effective, showing a smaller gain and a progressive phase lead
(Carpenter 1988). Our main hypothesis was that the otoliths would enhance the
gain and reduce the phase lead of the response in the lower end of the frequency
range.
In a previous study we measured
ocular torsion during sinusoidal body roll at 0.25 Hz about an earth-horizontal
and about an earth-vertical axis, and observed that the torsional nystagmus
contained less anti-compensatory saccades in the former condition, which corresponded
to larger amplitudes of ocular torsion (Groen et al. 1996b). A similar
finding has been reported for the vertical VOR in rabbits (Barmack 1981). We
believe that modulation of the saccadic activity in vestibular nystagmus offers
the possibility for the otoliths to control the position of the eyes in space,
and that this may be independent of effects on the slow component (velocity)
of the response. In addition to the slow component dynamics, we therefore investigated
the saccadic activity and the amplitude of torsional eye position in the original
nystagmus.
Methods
Subjects
Each of five healthy subjects, one female and four male,
28-37 years old, participated in one experimental session of an hour with their
informed consent. Seated in a rotating chair, the subjects were sinusoidally
rotated about their naso-occipital (roll) axis. The subjects� head was supported
by a head rest, and the body was firmly harnessed to the chair.
Stimuli
In total, there were six different stimuli of sinusoidal
roll motion. Four of them varied in frequency (0.05, 0.1, 0.2, and 0.4 Hz),
but had a fixed amplitude (25�), allowing for evaluation of the frequency response.
Moreover, at the frequency of 0.2 Hz there were two additional stimuli with
an amplitude of 12.5� and 50�, respectively, so that the linearity of the response
could be examined. Responses were measured during ten cycles at most, with a
maximum duration of one minute (that is 3 cycles at 0.05 Hz, 6 cycles at 0.1
Hz, and 10 cycles at 0.2 and 0.4 Hz).
The same set of stimuli was applied in two different
body orientations (Figure 3.1). In upright position, the rotation axis was earth-horizontal.
Here, the stimulus modulated semicircular canal and otolith inputs. In supine
position, the rotation axis was earth-vertical. Here, only the canal input was
modulated. Thus, there were 12 different conditions (six motion stimuli in two
body orientations). To save time of rearranging the chair between the two orientations,
conditions were not randomly applied. Instead, the stimuli were divided into
two blocks of three, each of which was carried out in the upright orientation
first, followed by the same block in the supine orientation.
Eye movement recording
With two small synchronized CCD-cameras mounted on a head
set, movements of both eyes were simultaneously recorded on video with a sample
frequency of 50 Hz. To avoid artifacts from movement of the cameras relative
to the head, the head set was attached to a dental frame with individual fit.
Except for a point source fixed to the camera, the measurements were performed
in darkness. The subjects were asked to gaze straight ahead into the cameras,
using a reflection spot on the camera lens for fixation. Thus "pure"
torsional movements were measured. The video recordings were digitized and analyzed
off-line by means of the method described in Chapter 1. The angle of ocular
torsion was determined relative to a reference image, arbitrarily chosen from
the recording just prior to the stimulus onset. The eye position signals were
stored in separate datafiles, synchronized with the corresponding stimulus signals
(the actual rotation angle of the chair).
Analysis of the slow component
The datafiles were analyzed using an interactive computer
program. First, we determined gain and phase of the slow component velocity
(SCV). For this, the position signal was differentiated digitally, and saccades
were removed by a moving median filter with a window of 500 ms (the saccades
were typically 100 ms of duration, or less). Subsequently, the amplitude and
phase of the component with the stimulus periodicity were estimated by means
of discrete Fourier transformation. The SCV gain was computed as the amplitude
ratio of eye velocity and stimulus velocity. The SCV phase was determined relative
to the stimulus. Positive values were used for a phase lead, where zero phase
indicated perfect compensatory eye velocity.
Although the gain and phase are interdependent parameters,
we examined them separately. This implies that an ambiguous phase of a small
response may have a disproportionate weight in the group�s average. Anticipating
the results, however, no data had to be excluded for this reason. First, all
individual responses were distinct enough to allow for reliable parameter estimation.
And second, the between-subject variability was small.
Analysis of eye position
In the absence of saccades, the behavior of the eye position
would be directly described by the SCV parameters. But in any nystagmus the
excursion range of eye position is modified by saccades. To examine the effects
of saccades, we therefore determined the gain of torsional eye position, by
averaging a number of cycles of theoriginal nystagmus (including saccades) and
measuring the amplitude of the approximately sinusoidal result. Analogous to
the determination of the SCV gain, this amplitude then was divided by the stimulus
amplitude to obtain the eye position gain.
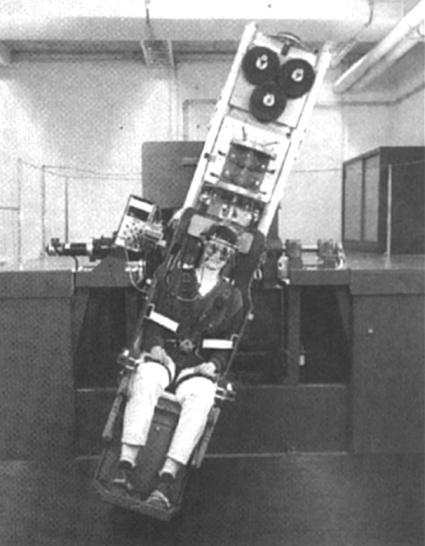
Figure 3.1. Two pictures showing the orientation of the TNO tilt chair for
whole body roll about (a) an earth-horizontal axis and about (b) an earth-vertical
axis. To illustrate the camera equipment for OT recordings, the hood was removed
in (a). The roll axis was more or less centered between the eyes.
The saccades themselves were characterized
by their amplitude and incidence (frequency). In general, the saccade amplitude
is monotonously increasing with peak velocity (Bahil et al. 1975, Lebedev
et al. 1996), a rule which also holds for torsional saccades (Collewijn
et al. 1985). For technical reasons we took the saccade peak velocity
as a measure for their amplitude. The saccade frequency was expressed
as the number of saccades per second.
In general, the above analyses
were performed on the entire response, with disregard of the first cycle to
allow the response to build up. Incidentally, one or more cycles had to be ignored
because of artifacts (e.g. eye blinks).
Hypothetical otolith response
Assuming additive contributions of the semicircular canals
and the otoliths to the slow component, we calculated a hypothetical otolith
response as the difference between the upright and the supine response. Instead
of actually subtracting the original signals, we calculated averages of the
individual vector differences using the available gains and phases of the SCV.
Results
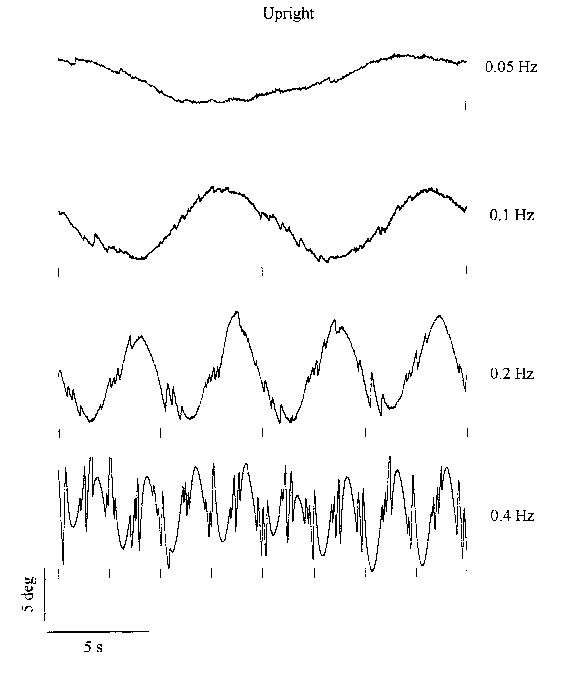
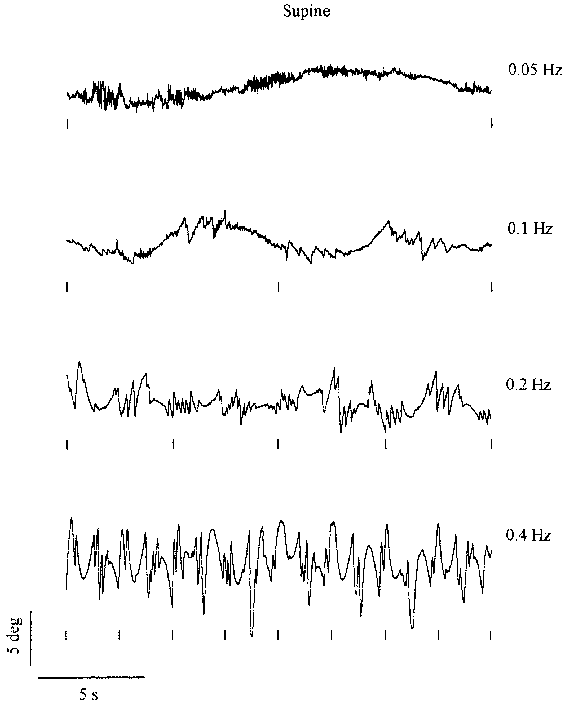
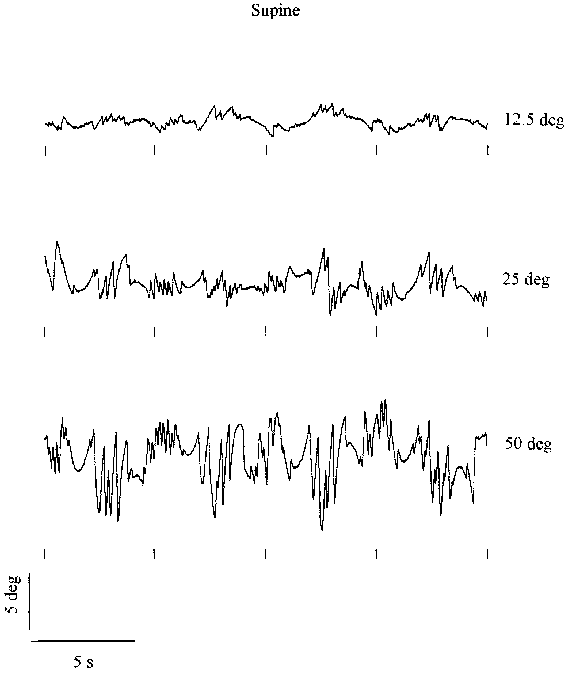
Figure 3.2 shows 20 s periods of torsional nystagmus of
one subject at different stimulus frequencies, with a fixed amplitude of 25�.
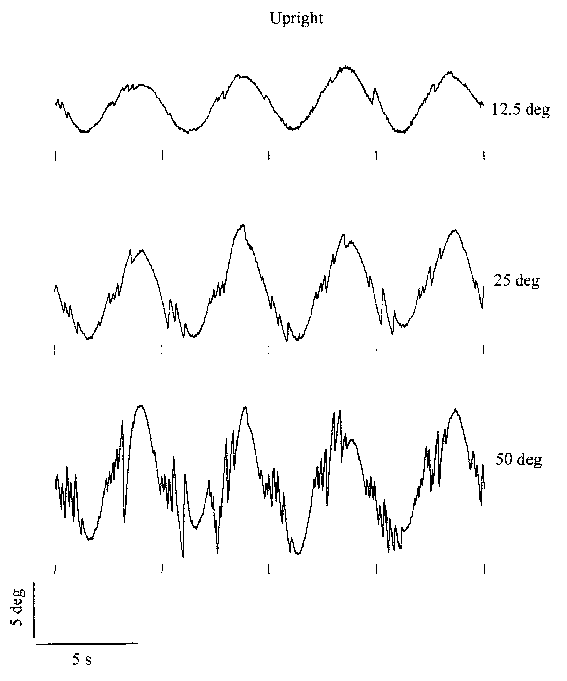
Data are shown in both the upright and supine conditions. Similarly, Figure
3.3 shows data of the same subject at different stimulus amplitudes, where the
frequency is fixed at 0.2 Hz. All responses feature an obvious sinusoidal "slow"
component sharing the periodicity of the stimulus. The amplitude of the nystagmus
appears to increase with increasing stimulus frequency and amplitude. In the
next paragraphs the contribution of the slow component and the saccades will
be quantified.
Slow component velocity
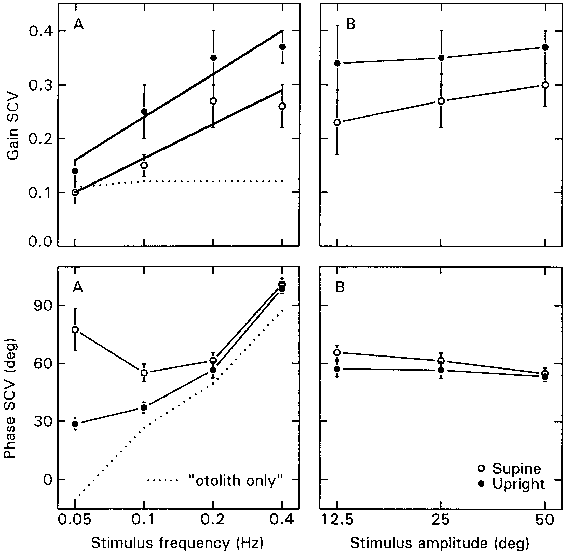
In Figure 3.4a the SCV dynamics (gain
and phase), averaged over all subjects, are plotted versus stimulus frequency
(with an amplitude of 25° ). The data of both eyes have been combined.
The SCV gain increases significantly with stimulus frequency (within subjects
design ANOVA: F=34.87; df=3,12; p<0.01). Furthermore, the average gain is
significantly higher in upright than in supine orientation (F=29.80; df=1,4;
p<0.01). The linear relationship between the SCV gain and the logarithm of
the stimulus frequency (f) is demonstrated by the two regression lines, plotted
in the upper diagram of Figure 3a: yupright = 0.50 + 0.26 log(f),
and ysupine = 0.37 + 0.21 log(f).
The corresponding phase characteristics,
as presented in the lower diagram of Figure 3.4a, reveal a significant interaction
between body orientation and stimulus frequency (F=11.07;df=3,12; p<0.01).
In the supine orientation, at 0.05 Hz the response leads the stimulus on average
by 78�, which decreases slightly at 0.1 Hz, and increases again up to 101� at
0.4 Hz. In the upright orientation, the average phase lead is only 29� at 0.05
Hz, and successively increases to 99� at 0.4 Hz. From 0.2 Hz and above, the
phase leads in both body orientations are identical.
The hypothetical "otolith only"
response, the vector difference of the upright and supine response, is also
depicted in Figure 3.4a (dotted line). The gain of this response is about 0.1
for all frequencies, where the phase changes from a lag of about 15� at 0.05
Hz into a lead of 90� at 0.4 Hz.
In Fig. 3.4b, the SCV gain and
phase are plotted versus stimulus amplitude (with frequency fixed at 0.2 Hz).
There was no effect of amplitude, which implies that the response behaves linearly
in both body orientations for amplitudes from 12.5� to 50�. Again, there is
a main effect of body orientation on the SCV gain, which is consistently higher
in upright than in supine (F=59.87;df=1,4;p<0.01).
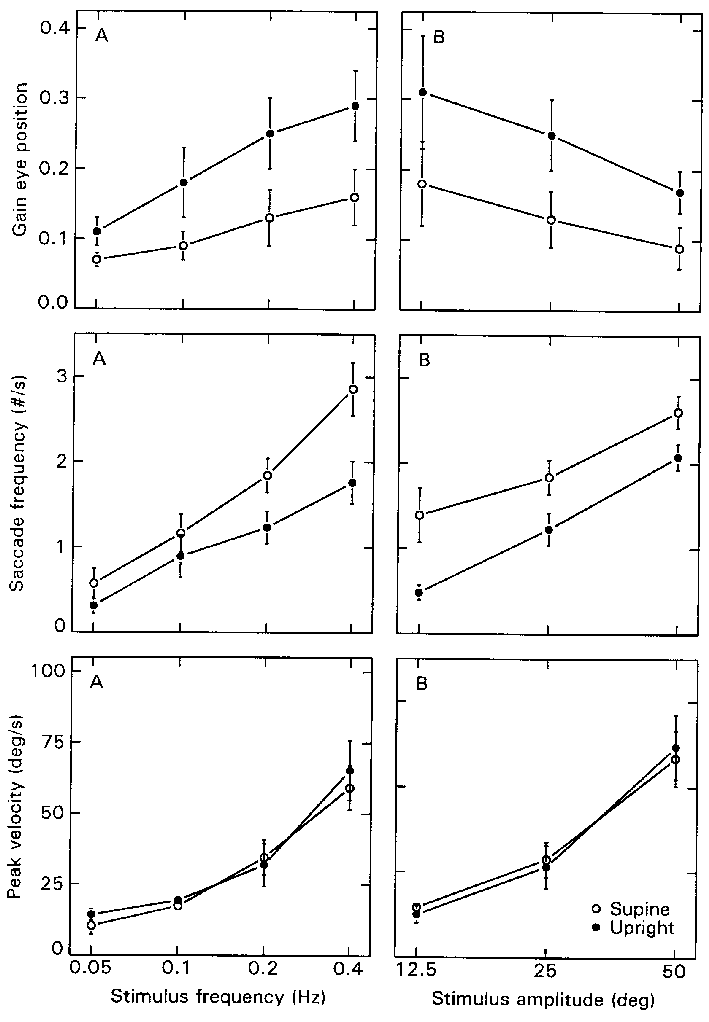
Eye position
The three parameters describing eye position behavior are
plotted in Fig. 3.5a for increasing stimulus frequency, and in Fig. 3.5b for
increasing stimulus amplitude. The excursions of torsional eye position are
invariably smaller than the corresponding SCV gains. This confirms that the
saccades are oppositedly directed to the slow component, reducing the maximum
amplitude of torsion. This is best seen by comparing the upper diagrams of Figures
3.4b and 3.5b: the eye position gain decreases significantly (F=64.78;df=1,4;p<0.01),
whereas the gain of the SCV remains constant with increasing stimulus amplitude.
Parallel to this, the average saccade frequency increases significantly with
stimulus amplitude (F=15.76;df=2,8; p<0.01), as does the peak velocity (F=49.84;df=2,8;
p<0.01). Similarly, the saccade frequency increases with stimulus frequency
(F=29.28; df=3,12; p<0.01), as does the peak velocity (F=23.24; df=3,12;
p<0.01). In this case, however, the gain of the eye position still increases,
due to the increase in SCV gain. From the original recordings, as shown in Figures
3.2 and 3.3, it is evident that the saccadic activity is higher supine than
upright. In fact, the saccades seem to operate around an "offset",
which appears sinusoidally modulated in the upright conditions, but seems about
zero in the supine conditions. Figure 3.5 shows that this can be attributed
to a difference in the saccade frequency, which is significantly higher in the
supine than in upright orientation (F=26.76; df=1,4; p<0.01). On the other
hand, the saccade amplitude (or "beating field") is independent of
body orientation, as is indicated by the average peak velocity, which does not
differ between upright and supine conditions.
Figure 3.2. Twenty seconds of torsional nystagmus of one subject at four
stimulus frequencies (fixed amplitude 25 �) in the upright (left) and supine
(right) orientation. The small markers below each trace indicate the moment
of maximum clockwise body tilt. Upward deflection of the eye position signal
corresponds to counter-clockwise rotation. (Click graphs to enlarge)
Figure 3.3.
Twenty seconds of torsional nystagmus at three different stimulus amplitudes
(with frequency fixed at 0.2Hz) in upright (left) and supine (right). Data are
from the same subject as in Figure 3.2. The small markers below the traces correspond
to maximum clockwise body tilt. (Click graphs to enlarge)
Figure 3.4.
The SCV dynamics, binocular data averaged over five subjects, for the upright
(filled circles) and supine conditions (open circles). In the left column (a)
the SCV gain and phase are depicted versus log stimulus frequency. Here, the
dotted line shows the hypothetical "otolith only" response, calculated
as the vector difference between upright and supine. The solid lines in the
gain plot (upper left diagram) are the fitted regression lines: yupright
= 0.50 + 0.26 log(f), and ysupine = 0.37 + 0.21 log(f). In all other
plots the connecting lines between data points are drawn. In the right column
(b) gain and phase are shown for the log stimulus amplitude. Positive phase
values indicate phase lead relative to a perfect compensatory response. Error
bars indicate S.E.M.
Figure 3.5.
The eye position parameters as they vary with log stimulus frequency (a, left
column) and amplitude (b, right column). The mean eye position gain (upper diagrams),
saccade frequency (middle diagrams), and saccade peak velocity (lower diagrams)
are shown for upright (filled circles) and supine conditions (open circles).
Error bars indicate S.E.M.
Discussion
In this study we compared the torsional VOR during oscillations
about an earth-horizontal and an earth-vertical roll axis, that is with varying
and invariant otolith input, respectively. We used passive body roll in order
to minimize inputs from the neck receptors. Therefore, the response differences
between the two body orientations were attributed to the contribution of the
otoliths.
Slow component velocity
We found that in both body orientations
the gain of the slow component velocity increased with stimulus frequency. The
gains observed were within the common range of the human torsional VOR. For
instance, the average SCV gain of 0.37 at 0.4Hz in the upright condition is
similar to the value measured by Tweed et al. (1994) at 0.3Hz. A further
increase of the torsional gain up to 0.7 has been reported for active head movements
of 1 Hz (Collewijn et al. 1985; Vieville and Masse 1987). The logarithmic
relation between the SCV gain and the stimulus frequency, suggested by our data,
confirms these results.
In the present study, the gain
was consistently higher, by an amount of 0.1, in the upright orientation than
in the supine orientation. In a previous study we measured the torsional response
in exactly the same way at the frequency of 0.25 Hz (Groen et al. 1996b).
We then observed a higher gain in the supine conditions compared to the upright
conditions. Among the 11 subjects of the previous experiment were the same five
subjects of the present study. Although the difference between the two conditions
was significant for the entire group of 11 subjects, it was not for the subgroup
of five subjects. Later we collected more (unpublished) data of the same subjects,
again at the frequency of 0.25 Hz, which were consistent with the results described
here. Considering these variable results within subjects and the small effects,
we do not believe that the otoliths contribute significantly to improve the
gain of the (torsional) VOR in humans. Other studies have also failed
to show a difference in SCV gain between earth-horizontal and earth-vertical
rotation (Tweed et al. 1994; Seidman et al. 1995). In this, humans
differ from lateral-eyed species, such as rabbits, where the otoliths almost
completely compensate for the reduced gain of the canal-induced VOR at very
low stimulus frequencies (Barmack 1981).
We adhere to a different conclusion
with respect to the phase of the response: in the lower frequency range,
contribution of the otoliths had a significant effect on the phase lead. At
the frequency of 0.05 Hz the response to earth-vertical rotation showed a phase
lead of about 80�, which was only 30° in the response to earth-horizontal
rotation. Similarly, Vieville and Masse (17) reported that the phase lead of
ocular torsion during active head roll in their (only) subject was larger when
the head was prone than when the head was erect. There is evidence that otolithic
control of the phase is not restricted to the torsional VOR. The same effect
was reported for the phase of the horizontal VOR in humans during barbecue spit
rotation (Benson 1970). Optimizing the phase therefore seems to represent a
general mechanism of the otoliths to improve the VOR dynamics.
In the present study, there are
two otolith-induced responses under consideration: a (position-dependent) response
to head tilt, and a (position-independent) response to the rotation of
the gravito-inertial force relative to the head ("tilt velocity").
With our experimental design however, it is difficult to discriminate the two
components in the calculated "otolith only" response. This may explain
why this response did not show the typical low-pass filter characteristics,
which have been described for tilt OORs (Paige and Tomko 1991a; Hannen et
al. 1966; Lichtenberg et al. 1982; De Graaf et al. 1996; Merfeld
et al. 1996). It is possible that the observed otolith effect on the
phase of the slow component exclusively resulted from the response to tilt velocity,
as was argued by Angelaki and Hess (1996a). Using sinusoidal rotation with an
amplitude of more than 360�, these authors demonstrated that it was the "tilt
velocity" information by which the otoliths improved the low-frequency
dynamics of the angular VOR in monkeys.
Theoretically, the reduction
of the phase lead of the slow component may be accomplished via the neural integrator
(Robinson 1971), which converts eye velocity signals into position signals.
In their physiological range, the canal afferents encode for angular head velocity,
which is suitable to drive the movements of the eyes, but needs to be integrated
to hold the eyes in their new position after cessation of the movement. At lower
frequencies, the canal signal is shifted toward acceleration (Férnandez
and Goldberg 1971), as becomes clear in the progressive phase lead of the VOR.
To hold eye position then, the signal would have to be integrated twice. Our
data suggest that the otoliths provide supplementary (tilt) velocity information
that can be used to achieve this extra integration step.
Eye position
Due to anti-compensatory saccades the eye position gain
was invariably smaller than the gain of the slow component. This effect was
most noticeable in the response to earth-vertical rotation: at all stimulus
frequencies the nystagmus consisted of more saccades in the supine condition
than in the upright condition, even though the corresponding SCV gain was higher
in the latter. The amplitude of the saccades was equally large in both body
orientations, indicating that the beating field itself was unaffected. Instead,
it was the offset of the beating field that was sinusoidally modulated
during earth-horizontal rotation. This resulted in larger accumulation of ocular
torsion in the upright orientation, illustrated by the higher eye position gain.
This confirmed our assumption that the otoliths modulate torsional eye position
by influencing the saccadic behavior. In contrast to the observed effect on
the phase of the slow component, the effect on the saccade frequency reached
over the entire frequency range, so that presumably the two effects are independent.
We believe that modulation of the beating field offset
is an instrument for the otoliths to maintain eye position in space. It may
well be that this is common to any system associated with gaze stabilization,
since it has been reported that the incidence of anti-compensatory saccades
in the ocular torsion response is reduced in the presence of a visual scene
(Collewijn et al. 1985). By the same token, our results indicate that
stabilization of the eyes-in-space is less important when the rotation is independent
of gravity. Here, the system seems inclined to compensate for head velocity,
irrespective of the orientation of the eyes. From this we conclude that the
canals are primarily concerned with stabilization of the retinal image in head-centric
coordinates, whereas the otoliths tend to anchor the oculomotor coordinate system
with respect to gravity.
Listing�s law
Although our study concerned one-dimensional eye movements,
the above described effects are relevant in the light of Listing�s law, which
applies to three-dimensional eye movements. According to this law, the eyes
usually assume positions that can be reached from a certain reference position
by rotation about an axis laying in a head-fixed plane. When this reference
position is the primary position, these axes lie in a plane (Listing�s plane)
perpendicular to the gaze direction (Tweed et al. 1990). As a result
of the strategy as described by Listing�s law, the oculomotor system tries to
avoid torsional eye positions. This implies that there is a conflict between
minimizing the degree of ocular torsion and stabilizing the position of the
eyes in space. This raises an alternative explanation for the low torsional
gain of the VOR. Originally, it has been argued that a low torsional gain is
not necessarily inadequate, because torsional movements do not alter the direction
of gaze and thus preserve fixation on a stationary target (Seidman et al.
1989; Ferman et al. 1987). As a result, there would be little retinal
slip in the foveal area. However, these assumptions only hold when ocular torsion
is regarded as rotation about the line of sight, that is relative to the eye.
There is reason to believe that the torsional VOR should be regarded in head-fixed
coordinates, or, more precisely, in Listing�s coordinates (Crawford and Vilis
1991; Tweed et al. 1994). In the view of these authors, the low torsional
gain signifies a trade-off between perfect (co-axial) compensation for head
rotations and obeying Listing�s law. The compensatory slow component tilts the
rotation axes of the eyes out of Listing�s plane, which is subsequently restored
by anti-compensatory saccades. Our data confirm this behavior for the supine
position. During earth-horizontal rotation, however, the (now) varying otolith
contribution complicates the matter. It is questionable whether this otolith-induced
ocular torsion should be considered as a violation of Listing�s law. In fact,
it may establish the link between the eye in its head-centric orientation (according
to Listing�s law) and the eye in the orientation to the environment (i.e. to
gravity). It has been hypothesized that the static ocular torsion during head
tilt may result from a shift of Listing�s plane (Vilis 1993). Indeed, the rotation
axes of voluntary saccades have been shown to lie in a plane shifted in torsional
direction when the subject is tilted laterally (Crawford and Vilis 1991). Similar
observations have been made for static tilt in pitch (Haslwanter et al. 1992).
Recently, Angelaki and Hess (1996a,b) found that in monkeys the otoliths did
not only influence the slow component but also the saccades of vestibular nystagmus.
They argued that the orientation of primary position is dynamically modulated
by the otoliths. The modulation of the beating field offset, observed in the
present study, indicates that the otoliths serve this function also in humans.
In conclusion, our data showed that the dynamic contribution
of the otoliths improved the human torsional VOR dynamics at low stimulus frequencies,
mainly by keeping the response in phase with the stimulus motion. In addition,
the otoliths reduced the number of anti-compensatory saccades in the nystagmus,
increasing the excursion range of torsional eye position. Through these effects
the otoliths achieve better stabilization of the eyes-in-space.
Acknowledgments
This work was supported by the Foundation for Behavioral
and Educational Sciences of the Netherlands Organization for Scientific Research.
The dental frames with individual fit were provided by ACTA (Academisch Centrum
Tandheelkunde Amsterdam). We are grateful to Astrid de Vries for her contribution
to the data analysis. We also thank Dr. Doug Crawford for his helpful discussions.
"Those who are to participate in the apparent gyrations of the swing -
and there may be quite a number who enjoy it simultaneously - are ushered into
a small room. From a bar crossing the room, near the ceiling, hangs a large
swing, which is provided with seats for a number of people. After the people
have taken their places, the attendant pushes the car and it starts into oscillation
like any other swing. The room door is closed. Gradually those in it feel after
three to four movements that their swing is going rather high, but it is not
at all. The apparent amplitude of the oscillations increases more and more,
until presently the whole swing seems to whirl completely over, describing a
full circle about the bar on which it hangs. To make the thing more utterly
mysterious, the bar is bent crank fashion, so that it seems demonstrably impossible
for the swing to pass between bar and ceiling. It continues apparently to go
round and round this way, imparting most weird sensations to the occupants,
until its movements begin gradually to cease and the complete rotation is succeeded
by the usual back and forth swinging, and in a few seconds, as the children
say, "the old cat dies." The door of the room is opened and the swinging
party leaves. Those who have tried it say the sensation is most peculiar and
the deception perfect."
"The haunted swing"
(from A.A. Hopkins. 1976. Magic, stage illusions , special effects and trick
photography. A.A. Hopkins. Dover Publications, New York)