Chapter 6
Altered behaviour of hamsters by prolonged hyper-gravity; adaptation to 2.5 G and re-adaptation to 1 G
Published in: Acta Otolaryngologica (Stockholm). 116:192-197, 1996
Abstract
We studied the functional adaptation process
in 40 hamsters subjected to either prolonged hypergravity or normal gravity.
Subadult golden hamsters (n=20) exposed to a hypergravity condition of 2.5 G
for 6 months were tested to investigate the effect of hypergravity on the perceptive-motor
skills and compared to control hamsters (n=20). The motor coordination of the
hypergravity hamsters hardly changed; locomotion was normal and swimming was
possible. Equilibrium maintenance was disturbed during the first 3 months as
was shown by the higher crossing time (p<0.001) and higher fall frequency
(p<0.001) for the hypergravity group. Significant differences were also found
in orientation during swimming (p=0.007) and turning behaviour in the rotation
task (p<0.001) and in the no-rotation task (p=0.029). After 6 months, 10
hamsters of both groups were tested for another 4 months, also the hypergravity
hamsters were living at 1 G. Differences in orientation in the two groups did
not change during swimming and turning behaviour during the rotation task (p=0.026).
Based on our findings, we conclude that
the hamsters functionally adapted to hypergravity, which led to an altered performance
of several tasks. The condition continued after 4 months of normal gravity.
Keywords: sustained acceleration, equilibrium maintenance, swimming behaviour, locomotor activity, animal experiments.
Introduction
The vestibular system is essential for optimal
spatial orientation, as alterations in the system affect stability in gaze,
posture and locomotion (Dai et al. 1994). In animal experiments, peripheral
vestibular functioning is investigated by tests directed to vestibular-ocular
reflexes, head movements, locomotion and swimming behaviour. For example, rodents
with artificial lesions in the otoconial layers or bilaterally labyrinthectomized,
show abnormalities in swimming, locomotion and air-righting (Huygen et al. 1986;
Ossenkopp et al. 1986; Llorens et al. 1993).
Prolonged increased gravity, hypergravity (HG), in
a centrifuge provides opportunities for research on the adaptation of sensory-motor
processes to a changed gravity condition. In earlier experiments, we found that
hamsters subjected to HG for 3 months, were less able to balance on tubes and
to orientate in a watermaze than control hamsters living under 1 G conditions
(Sondag et al. 1995a). This outcome suggests involvement of the vestibular system.
The aim of the present study was to investigate (i)
the functional adaptation process in hamsters exposed to HG for 6 months and
(ii) the re-adaptation, for another 4 months, under normal gravity conditions.
Material and Methods
A group of 40 male subadult golden hamsters
(Mesocricetus auratus, Harlan, Zeist, The Netherlands), with a mean weight
of 47 g were born and raised in normal gravity. After weaning, 20 hamsters were
placed in acrylate boxes (22 cm x 37 cm x 30 cm; 3 or 4 animals per box), inside
the centrifuge-gondola to live under conditions of 2.5 G (HG hamsters). The
20 controls (CON hamsters) were placed in similar housings under normal gravity.
Food and water were available ad libitum and the day-night cycle (light
on 19.00 - 7.00 h) was reversed. Testing started after 1 week of habituation.
The tests were done in a laboratory room with dimmed lights and additional noise
(radio) to avoid distraction of the hamsters by the experimenter. After 6 months,
10 of both groups were sacrificed (randomly selected) for histological and morphological
examination of the otoconia. The results of the latter study have been published
previously (Sondag et al. 1995b). The remaining 20 hamsters lived under 1 G
conditions for another 4 months. The body weight was measured twice a week.
The experiments were performed in accordance with the recommendations provided
in a special license as required by the Dutch Law on the Use of Animals in Scientific
Research.
The centrifuge consisted of a centrally placed 3.5-kW
DC motor drive and two horizontally mounted arms (length = 115 cm) with aerated
and darkened free-swinging gondolas (length 110 cm, width 45 cm, height 80 cm,
length arm + gondola 194 cm). A rotation speed of 34.3 rpm produced a 2.5 Gvalue
at the floor of the gondola (Sondag et al. 1995a).
Depending on the kind of task, tests were done weekly,
bimonthly or monthly. We registered the tubes and swimming (lane and maze) weekly;
the treadmill activity bimonthly; the rotation and the no-rotation task monthly
and the open field study once in two months.
Locomotion
To gain insight into the locomotor activity
of HG hamsters during centrifugation, a treadmill was placed in one of the boxes.
Daily, the treadmill activity of one hamster was recorded by means of a cyclocomputer
(CC-MT200, Cateye, Osaka, Japan) providing the average speed, the running time
and the distance covered in one day.
Open field study: each hamster was placed in the middle
of the open square (140 x 140 cm, height of the walls 30 cm) for 5 minutes.
The floor was cleaned before placing the hamsters in the square. The animal
behaviour was recorded on tape for further analysis. The floor surface was divided
in 196 squares of 100 cm2 (52 peripheral and 144 central squares).
We measured the traversed distance, grooming activity, number of faecal boli
an index of emotional reactivity), number of rearings (rearing up on hind legs)
and duration of each rearing.
Tube tasks; the acrylate tubes (length 100 cm, diameter 20
mm), placed ± 20 cm above ground level, were either fixed to standards (fixed
tube task) or were movable connected by elastic cords which were attached to
the standards (mobile tube task). Both tubes were covered with tape in order
to give the hamster a better grip. A platform was placed at the end of the tube
where the hamster could collect sun-flower seeds for ± 5 s. One extra day was
used to train the hamsters for 5 minutes to walk the full length of the tube.
Each hamster had to cross the tube 3 times (trials a, b, c). Tests were done
once a week. After one week of HG, testing on the fixed tube started. After
two weeks, testing on the mobile tube started.
Swimming behaviour and susceptibility for accelerations
Swimming speed in a straight lane (140 x 10 cm, height of the walls 30 cm, water-depth 25 cm, water temperature ± 300 C) to test swimming ability and speed. The hamsters had to swim to the end of the lane where they could climb an escape ladder. One extra day was used to train the hamsters for 5 minutes to swim to the ladder. On the testing days, the animals swam 6 trials; 3 in the lane and 3 in the watermaze. The crossing time for the middle part of the lane (length 100 cm) was measured.
Orientation ability during swimming in a
maze (140 x 70 cm). The shape of the maze was similar to our earlier experiments
(Sondag et al. 1995a). The hamsters had to find a ladder, located at the opposite
end of the maze. The crossing time was measured and the swimming strategy was
observed.
Rotation task; a hamster was placed in a cylindrical bucket
(diameter 30 cm, height 45 cm) mounted on a swivel tool device with 10 sinusoidal
pendular rotations at a frequency of 0.1 Hz and an amplitude of 900
(total swing time 100 s). The behaviour of each animal was recorded by a video-recorder
(SONY handycam) mounted at the top of the device.
No-rotation task; the hamsters' behaviour was also recorded
for 100 s when the device did not move (the period between the rotation and
the no-rotation task was 2 weeks). The number of changes in walking direction
(turnings) was counted in both tasks.
Statistics: Data were statistically assessed (significance:
p<0.05) with the Chi-square test (orientation in the watermaze), repeated
analysis of variance (body weight, treadmill, swimming speed, rotation and no-rotation
task, open field study) and repeated analysis of variance with covariates (tube
tasks). We used the statistical software SPSS PC+ 5.0 for our analysis.
Results
The body weight increased in all 40 hamsters, the 20 HG hamsters grew slower than the CON hamsters did under 1 G. After 6 months, the difference in body weight of the two groups increased to ± 25.3% (mean HG group 112 g, mean CON group 151 g, F(1,38)= 121.53, p<0.001). After the return of the HG hamsters to 1 G, the body weight of the HG hamsters increased faster than that of the CON hamsters (F(13,234)=4.77, p<0.001): after 4 months of normal gravity the HG hamsters weighted less than the CON hamsters (mean HG group 139 g, mean CON group 166.8 g, not significant)
Locomotion
Treadmill activity; The HG hamsters walked a smaller distance (mean HG 2,45 km, mean CON 8.9 km, F(1,33) = 124.53, p<0.001), achieved a lower average speed (mean HG 0.99 km/h, mean CON 1.51 km/h, F(1,33)=158,66, p<0.001) and spent less time on the treadmill (mean HG 148 min, mean CON 347 min, F(1,33)= 83.63, p<0.001) than the CON hamsters. The differences between the groups decreased in the course of the experiment. Distance (mean HG 1.74 km, mean CON 0.75 km), speed (mean HG 1.05 km/h, mean CON 0.89 km/h) and running time (mean HG 95 min, mean CON 44 min) increased for the HG hamsters after returning to normal gravity, the distance (F(1,15)=4.69, p=0.047) was significantly higher for the HG hamsters than for the CON hamsters.
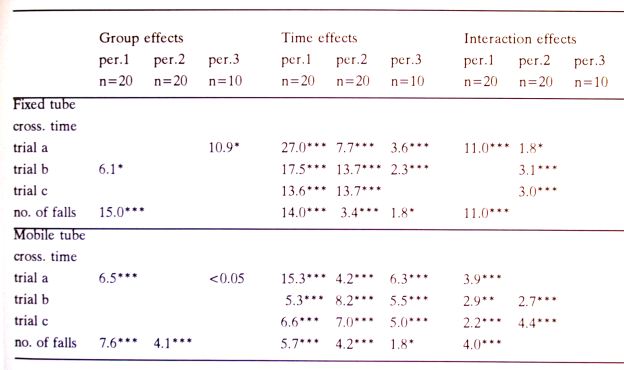
Table 1. Statistical results of the performance of the hamsters on the tube tasks during period 1, 2 and 3. Only significant F values and p-values (*p<0.05, **p<0.01, ***p<0.001) are given.
Open field study; The travelled distance decreased for both groups during the first part of the experiment (F(2,76)=7.85, p=0.001). No differences in travelled distance between the groups were found during the second part. The number of rearings increased in the first part (F(2.76)=4.54, p=0.014) but decreased in the second part (F(1,19)=7.66, p=0.012). The rearing time increased for both groups during the first part (F(2,76)=4.96, p=0.009) and was the same for the groups in the second part. No differences were found between the groups in grooming activity and the number of faecal boli.
For the tube and the water-maze task the differences in results between HG and controls disappeared in 3 months. Therefore, the data were analysed for 3 periods; the first part week 1 to 13, the second week 14 to 26 and part 3 week 27 to 40 (HG hamsters under 1 G conditions).
Equilibrium maintenance on the tubes
Fixed tube; After the first two weeks 10 of the 20 HG hamsters were not able to walk over the full length of the tube, the CON hamsters completed the three trials. After 2 weeks, all 40 hamsters finished three trials. Both groups learned to cross the tube in a shorter time during week 1 - 13 (period 1, Fig.1); the HG hamsters were slower to learn than the CON hamsters as was shown by the significant Group x Time interaction of the crossing time and fall frequency (Table I). During periods 2 (week 14 - 26) and 3 (week 27 - 40), the HG hamsters needed less crossing time than the CON hamsters. Differences in fall frequency between the groups were observed during the first trial of each testing day. During period 1, the fall rate of the HG hamsters decreased faster than that of the CON hamsters. In period 2 and 3, the fall frequency increased for both groups (Fig. 1, Table I).
Mobile tube; The data of the experiments were analysed in the same way as for the fixed tube. All 40 hamsters finished 3 trials on the first testing day. During period 1, the crossing time decreased slower for the HG hamsters than for the CON hamsters. In period 2, the crossing time increased less for the HG hamsters than for the CON hamsters. In period 3, no differences were found in crossing time between HG and CON hamsters. Differences in fall frequency were only observed in the first trial of each testing day. During period 1, the fall rate decreased slower for the HG hamsters. In period 2, the HG hamsters fell less often from the mobile tube than CON hamsters (Table I).
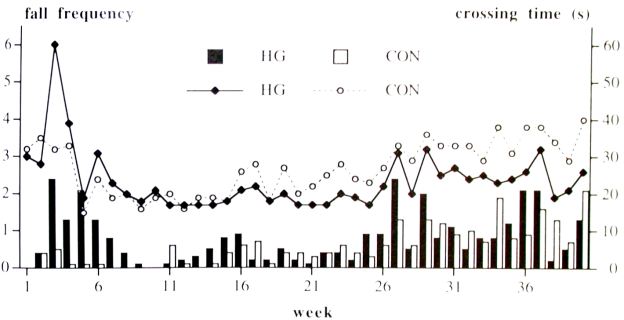
Fig. 1. Crossing time and fall frequency on the fixed tube. Bars represent the number of falls (Y1-axis), lines represent the crossing time (Y2-axis). Means (n=20 for week 1 - 26, n=10 for week 27 - 40).
Swimming behaviour and susceptibility for rotatory accelerations
Swimming ability; during period 1, the increase in swimming speed of the HG hamsters was lower than that of the CON hamsters (F(9,315)=2.99, p=0.02). In period 2 and 3, the swimming speed was the same for both groups (± 0.25 m/s).
Orientation in the S-maze; there were no differences in the crossing time between HG and CON hamsters. The crossing time of HG and CON hamsters decreased similarly for both groups in periods 1 (F(12,372)=7.53, p<0.001) and 2 (F(12,360)=1.83, p= 0.042) to ± 11 seconds for both groups at the end of period 2. Concerning the orientation strategy, both groups found the ladder by swimming along the walls of the basin during period 1. In week 14 - 25 (period 2), 5 HG hamsters (25%) and 11 CON hamsters (55%) swam straight to the ladder (p=0.007). The remaining 24 animals (15 HG and 9 CON) continued swimming along the walls of the maze. In period 3, each hamster kept its own strategy (straight to ladder: 2 of 10 HG (20%), 5 of 10 CON (50%).
Susceptibility for accelerations; during period 1 and 2, in the rotation (F(1,38)=56.13, p<0.001) and in the no-rotation task (F(1,36)=81.12, p=0.029) showed that the HG hamsters made less turnings than the CON hamsters. Moreover, in the rotation task, the CON hamsters increased the number of turnings, from 7.5 times at the start of period 1 to 12.8 times at the end of period 2 (F(1,190)=2.62, p=0.026). In period 3, the HG hamsters made less turnings than CON hamsters in the rotation task (F(1,15)=15.55, p=0.001; Fig. 2).
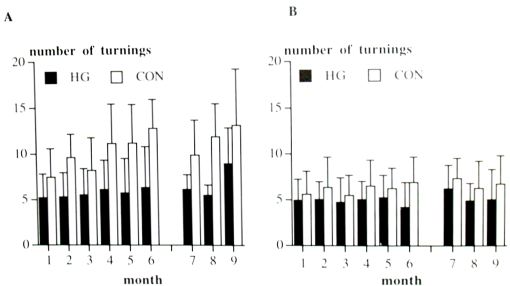
Fig. 2. Number of turnings during (a) the rotation task and (b) the no-rotation task. Means and standard deviations (n=20 for month 1 - 6, n=10 for months 7 - 9).
Discussion
We investigated the functional adaptation process in hamsters exposed to prolonged hypergravity. The HG hamsters gained less weight than the control hamsters and weighted less even after 4 months of normal gravity. Probably, this phenomenon was caused by an increased basal metabolism during the HG period (Economos et al. 1982).
The fact that the HG hamsters ran the treadmill shows they were motivated to perform locomotor activity other than eating and drinking. The activity level was lower for the HG hamsters than for the CON hamsters. We presume that this decrease in treadmill activity was caused by higher energy demands. The locomotor activity was the same in both groups in the open field, no differences between the groups were found in travelled distance, number of rearings, duration of each rearing, grooming activity and the number of faecal boli. This shows that the HG hamsters did not experience the transition from 2.5 G to 1 G as a stress full event and that locomotion became normal again when these hamsters were placed in the normal gravity environment with lower energy demands.
Compared to the CON hamsters, the HG hamsters had more difficulties in balancing on tubes during the first 3 months (period 1). For equilibrium maintenance, not only information from the vestibular system is needed, but also visual, proprioceptive and tactile information. We presume that the HG hamsters lacked appropriate vestibular information and learned to balance on the tubes by relying more on visual and tactile information (paws). Therefore, training increases the ability to walk on the tubes. During period 3 (at 1 G), the HG hamsters had fewer problems with balancing than the CON hamsters. Explanations for the decreased performance of the CON animals are the higher body weight and the increased body dimensions, making it more difficult to walk on small tubes.
The swimming ability in HG hamsters was not impaired; the speed in the lane was the same as for the CON hamsters, although the latter group reached the maximum speed earlier. The circling behaviour, seen by other investigators after artificial peripheral vestibular lesions (Huygen et al. 1986; Ossenkopp et al. 1992; Llorens et al. 1993), was not observed in our animals, thus showing that hypergravity caused less severe disturbances in the peripheral vestibular system. Our hamsters did not swim underwater as was observed in rats exposed to HG (Fox et al. 1992). However, those authors dropped the HG rats from 45 cm height in a supine position. Our hamsters were gently placed in the water and so resurfacing was avoided. In the watermaze, 11 of the 20 CON hamsters (55%) and only 5 of the 20 HG hamsters (25%) swam straight to the stair instead of swimming along the walls. When living under 1 G, the percentage did not increase for the HG hamsters (2 of 10). Visual, tactile and vestibular information are needed for spatial orientation (Douglas et al. 1979; Miller et al. 1982). Water immersion deprived the hamsters from somatosensory information. During spatial orientation in the watermaze, visual guidance has to compensate for the insufficient vestibular support in the HG hamsters. However, the visual system of hamsters is poorly developed (Finlay and Berian, 1985). Therefore, the target must be nearby for proper use of visual information. This condition can be realized by swimming along the walls.
In the rotation task, the HG hamsters made fewer turnings than CON hamsters and this number did not change and remained the same as in the no-rotation task, whereas the CON hamsters increased the number of turnings in the rotation task from 7.5 times at the start of period 1 to 12.8 times at the end of period 2. When moving around inside the centrifuge, the HG hamsters adapted to a compound of high linear and rotatory acceleration. We suggest that the HG hamsters were less sensitive to a moderated compound of accelerations under normal gravity than CON hamsters. This led to a weaker response in the rotation task, even after having lived in normal gravity for 4 months.
Based on our findings, we conclude that hamsters functionally adapt to a prolonged hypergravity condition. This leads to alterations in sensory motor processes during equilibrium maintenance, orientation while swimming and turning behaviour in the rotation task. The motor coordination of the HG hamsters was not seriously impaired because normal locomotion and swimming were still possible. Performance of tasks in which the animals can rely on other sensory input, such as proprioceptive or tactile information, could be improved by extra training as was seen in the tube tasks. With respect to spatial orientation, the results from the watermaze and the rotation task suggest that the vestibular sensitivity to normal gravity is decreased. During 4 months under normal gravity conditions, after the hypergravity period, these differences in orientation during swimming and turning behaviour remained, indicating a persisting behavioural adaptation to hypergravity during normal gravity.
A study on the structure and function of the peripheral vestibular system in animals conceived, born and raised under hypergravity conditions, is well underway. The results will be reported in the near future.
Acknowledgements
The authors gratefully acknowledged the Netherlands Organization for Scientific Research (NWO) for funding this project. This research was conducted while HNPM Sondag was supported by a grant of the Foundation for Behavioural and Educational Sciences (SGW) of this organization (575-62-049), awarded to Prof. Dr. WJ Oosterveld.