The role of the sensory cells
The sensory cell system is one of the organs of graviception. It is
highly sensitive to linear acceleration up to about 0.1 g (Ishay et al, 1989)
and to radial acceleration up to 4-5 g. As afore mentioned there are 3 types
of cilia with distinct morphology. Although the specific role of each is still
unclear at this point, we conjecture that the cilia at the base of the conus
and the paraocellar organ are responsible for the detection of linear-horizontal
and vertical accelerations. While the cilia on the three periocellar crests
probably are responsible for the detection of radial accelerations in all three
spatial axes. The sensory information from the "Ishay Organ" is integrated with
the input arriving from the peripheral photoreceptors of the frons and vertex.
This integration takes place in a special ganglion, near the "Ishay Organ" and
is transmitted to the protocerebrum to be further integrated with information
coming from the 3 ocelli.
Light absorption by compound eyes and ocelli
As for visual organs intended to absorb light, there are three types, namely,
compound eyes (2 of each), ocelli (3 of each) and a large number of extraretinal
photoreceptors. The last ones are scattered, both on various regions of the
head, as well as on the surface of the body. It is commonly believed that the
structure of the compound eye, the ommatidium, enables the detection of a complete
image and thus fulfills the normal function of vision. However, the ocellus
is constructed to pick up polarized light and also enables seeing in the dark.
So, in hornets the compound eye handles the visual needs, while the ocelli serve
to locate the direction of the light source and hence is of importance in navigation
and orientation. The sensing of dim light and polarized light is also important
in the nest where there is relative darkness day and night. Sources of light
in the nest are the nest entrance during daytime, the yellow pigment in the
adults cuticle, yellow pigment in the larva's head and the silk of the cocoons,
all of which are luminescent and active day and night. In the nest the ocelli
might serve for orientation towards the nest opening, for communication or for
various nest activities.
Micromorphology of the ocelli
Further investigations on the micromorphology of the ocellus show that
the ocellus is composed of numerous functional units. However, essentially each
visual unit in the ocellus is comprised of two retinular cells which are joined
together at the base by a common rhabdom. The presence of the rhabdom in depth
enables vespan sensing polarized light and dim light. In fact, it is a structure
typical of a scotopic eye. The dispersal of the pigments (melanin) within the
retinular cells promotes the blocking of each unit (in the ocellus) from nonincident
light and ensures that each ocellus will respond only to directly incident light.
On the other hand, in the compound eye there is a short conus which transmits
light in the direction of the rhabdom, it is composed of an arrangement of 7
retinular cells. Here, the pigment granules are distributed at the bases of
the rhabdomeres. Structurally, this can be compared with a photopic eye. Another
difference between the simple and compound eyes is that each ommatidium has
its own separate cornea, whereas all the ocelli share their cornea. Both, the
compound eye and the ocelli are comprised of numerous autonomous units, each
of which gives rise to a separate axon. These axons subsequently join to form
the optic nerves, one for each compound eye, and one for each ocellus and these
ultimately are connected with the protocerebrum.
Arrangement of the Ocelli in the Hornet's Head
The ocelli are situated on the vertex plate in such a fashion that if we draw
a straight line through each, we end up with an equilateral triangle (Figs.
1-2). Each ocellus bears a convex cornea shaped as a hemisphere. The tangential
planes of each ocellus create in between a pyramid with three equal sides, as
shown in Fig. 1. The arrangement of each ocellus on a different plane enables
a panoramic visual coverage of practically all the 360 0 field of vision above
and around the head. On our stated assumption that the ocelli pick up polarized
light, we surmise that hornets can sense the direction of the sun rays through
them. However, to be able to determine the direction and angle of the sun vis-a-vis
the zenith which is antipodal to gravity, the hornet needs to orient itself
in a position which is absolutely tangential to the earth's surface. For this
purpose, the hornet relies on two organs of equilibrium located at some distance
from the two lateral ocelli, and a third, within the coronal suture which extends
from the median ocellus. All these elements three are sensitive to linear acceleration.
The two paraocellar organs act like a level with a long arm and between them
is the coronal suture, thereby enhancing the sensitivity to every small imbalance
(on the part of gravity). With the aid of its equilibrium organs the hornet
locates the zenith, while placing itself during flight in a perfectly horizontal
position so that it can measure the angle of the sun in respect to the zenith.
By way of analogy, we could say that the hornet uses its organs of equilibrium
to calibrate its measuring device to the zenith.
Equilibrium and orientation
The described system is composed of 3 ocelli and 3 gravity receptors.
Such a system has to satisfy two main needs while in flight, namely, equilibrium
and orientation. As for equilibrium, there are three axes around which a flying
body can move, to wit: roll, pitch and yaw, where roll is the movement around
the longitudinal axis of the body, pitch is the movement around the transverse
axis of the body and yaw is the movement around the vertical axis. During hornet
flight, which almost invariably takes place in light, the roll movement is measured
by the change in the angle formed between the direction of the paraocellar organs
and the direction of the gravitational force, as well as by changes in the angle
of the conus, Fig. 1. The nerve fibers in the conus are orientated parallel
to the surface of the earth, while the bobs on the fibers are of course invariably
directed downwards, thus indicating the direction of the gravitational pull.
On viewing from the outside, what can be deemed a factual change, is that the
longitudinal axis of either the flying or walking hornet forms a right angle
(90 0 ) with the coronal suture on its frons (i. e. on its face). As for the
pitch movement, each change here results in a change in the orientation in the
vespan ocelli vis-a-vis the sun and also changes in the orientation of the paraocellar
organs and the paraocellar crest of the lateral ocelli. Finally, with regard
to roll movement, this induces changes in the angle between the ocelli and the
zenith and that between the gravitic organs and the gravitational force, so
a roll to the right causes the left paraocellar organ to orientate more in the
direction of gravitation. The right paraocellar organ moves away from the gravitational
pull, whereas a roll to the left has the opposite effect. At the same time,
the roll movement is sensed by the median paraocellar organ as well. As for
the yaw movement, it apparently helps to retain the existing angle between the
gravitic organs and the gravitational pull, while causing change in the angle
between the ocelli and the sun.
Do hornets have a gyro system ?
Is it possible to identify in the Oriental hornet a gyro system? I believe
so. Around each ocellus there is a curved, closed canal filled with a liquid,
probably hemolymph, and around each canal there is a crista, likewise circular,
the mouth of each crista bears a coating of cilia. In the course of rotational
movement, a difference is created in the acceleration of the contained hemolymphatic
fluid vis-a-vis the (solid) crista and its ciliary coating, which results in
bending of the cilia in a direction opposite to the rotational movement and
thereby to the incipience of radial acceleration.
Insofar, as each of the three crista-equipped ocelli is situated at an inclination
of 90 0 to the other two, it stands to reason that this configuration enables
simultaneously determination of the radial acceleration in each of the spatial
axes. In fact, each triad of ocellus, canal and crista comprises an independent,
selfcontained gyro system in which, during flight, the fixed point is situated
in the center of the ocellus and determines the linear acceleration of the gravitation
which is antipodal to the zenith. Together, all three ocelli encompass the 360
0 of spatial panorama and in this respect are analogous to the semicircular
canals on both sides of the head of vertebrates, except that in our hornets
each ocellus possesses a fully circular canal. The possession of a completely
circular canal, apparently, is more advantageous than that of a semicircular
canal for insect equilibrium during flight. For example, birds and bats can
veer slightly sideways during flight but do not roll over completely as do flying
insects attempting to attach to the ceiling. Even in the dark nest the gyro
system is apparently operative, mainly to determine the direction of gravity
during comb building. In the latter instance the fixed point is determined by
the conus in the "Ishay Organ" and the paraocellar organs which are oriented
toward the gravitic acceleration which is linear. During comb building, the
hornet circles the building site, probably in order to stir and activate the
fluid in the gyro system and thereby enable precise determination of the desired
direction of building.
Determination of the zenith
When the hornet walks in dim or dark surroundings it flips over so that
its dorsum and the caphalic ocelli are now on the underside and in this fashion
the hornet walks on the ceiling which constitutes the zenith. We suspect that
in this situation the bond between the ocelli and the gravitic organs is disrupted
leaving only the force of gravitation as the dominant factor. The latter are
engaged in determining that the nest direction will be toward the gravity detected
resultant, while the ocelli in this situation are engaged in other nest mates
by their luminescence. As for the hornet's assessment of the sun's angle vis-a-vis
the zenith, we note that any determination of the zenith is made in the course
of rotational movement during ascending flight in the open air. The rate of
this rotational movement is at first slow, but it gradually accelerates till
the resultant is at the appropriate angle with respect to the angle between
the sun and the zenith. In this situation, the radial acceleration, which is
measured by the periocellar system, reflects the angle of the resultant vis-a-vis
the direction of the gravitational force, which is identical here to the angle
of the sun vis-a-vis the zenith. In this situation, also, both sides of the
median ocellus are equally lit while the illumination on the lateral ocelli
is symmetrical -all this provided the hornet faces the sun.
Orientation and ocelli illumination
When the hornet performs its orientation spin the sun illuminates the
ocelli at a varying angle which is dependent on two variables, to wit: (1) the
time of the day -in that the earlier or later the hour, the lower the sun's
position in the sky. This means that the more peripheral portions of the lateral
ocelli are illuminated along with the central portions of the median ocellus.
Whereas the closer the hour to noon is, the more the central portions of the
lateral ocelli are illuminated along with the peripheral portions of the median
ocellus. (2) the angle of the sun with respect to the zenith. For example, when
the sun is in its typical early noon position, its rays impinge on the hornet's
ocelli as the hornet performs its orientation rotatory movement which is counterclockwise
(vide lateral right ocellus in Fig. 3). However as the hornet's face is directed
towards the sun, point A on its right ocellus is illuminated, but as the hornet
continues its revolution the point of illumination passes directly through the
center of the ocellus to point B which is at 180° to point A. With continuation
of the hornet's rotatory movement to the point of solar illumination it passes
through points C and D before returning to point A upon completing one rotation.
What we get on the right ocellus, figuratively speaking, is a bow with a single
string, where in the angle between the string and an imaginary line transecting
the ocellus into an exterior and an interior half is precisely concordant to
the angle between the sun and the zenith. (Note that in Fig. 3, the lettering
pertaining to the ocelli refers either to the early and late hours -the capital
letters -or the noon hours -the small letters). Concurrently, movement of illumination
occurs also with regard to the left lateral ocellus, except that, now the ocellus
remains totally unilluminated throughout the movement from point A to point
B (i. e. half of a revolution). The pyramid of the vertex is shading the ocellus,
but it starts to be lit on point C and the illuminated point continuous to move
towards point D, and then from D back to A. From C to A, as well, one can draw
a hypotenuse whose angle vis-a-vis the transector of the ocellus will be congruent
with that between the sun and the zenith. Again, at the same time, a similar
motion occurs in the median ocellus, only now in the peripheral part of the
ocellus. Thus, as we proceed from A to B during the first quarter of the rotation
and from B t°C during the second quarter we observe illumination of the ocellus
but thereafter, during the concluding two quarters of the rotation, there is
no illumination of the ocellus. The angle between the hypothenuse drawn between
points D and B and an imaginary line transsecting the ocellus into front (bottom)
and back (top) halves is once more concordant with the angle between the sun
and the zenith. When the hornet makes its revolution clockwise, the situation
is reversed.
The "bee dance" equivalent
In Fig. 3, the path formed by sun rays on the ocelllus (right posterior
ocellus) is shown as a semicircle with a string in the center pointing to the
direction of the sun vis-a-vis the zenith. This circular (or actually semicircular)
path bears a resemblance to the circular (and actually semicircular as well)
path which a bee follows in the course of the know "bee dance", whereby the
dancing bee transmits information on the path traversed in the field in the
way for a food source, intended to reruit other bees to the task of collection
(Wilson, 1971). Of course, in the case of bees, where the combs are vertical,
as in the hive of the honeybee Apis mellifera, the relevant dance, named
the "waggle dance", translates the direction to or from the sun as the pivotal
pathway. In this way movements up or down vis-a- vis direction of the sun, are
interpreted as towards or away from gravity. When a bee 'dances' at an illuminated
site, like the mouth of the hive, or when bees build a horizontal comb, as Apis
florea does, the relevant dance also becomes a horizontal rather than a
vertical one. In hornets, whose comb is invariably horizontal and the cell outlets
face down toward the gravitational force, there is no dance as in bees, but
yet the sun rays impinge upon the ocelli in similar fashion. This leads us to
speculate that perhaps what the honeybee is actually reconstructing during its
dance in the hive proper (vertical) or at the hive entrance (horizontal) is
the trajectory of the sun rays upon its ocelli.
The "clock" of the hornet
Where, then, is the 'clock' of hornets located ? To the best of our
knowledge we have to seek for it in the ocelli, which to our mind are the timepiece
which lets the hornet know the time of day for as long as the sun is in the
sky. By way of explanation the following scenario is offered: Let us focus on
the orientation flight of the hornet, undertaken when it leaves the nest or
when it returns to the nest or, at times, also in the midst of regular flight.
At that time, the flying hornet faces the sun and in the morning hours, as the
hornet rotates counterclockwise, as observable in Fig. 4. Its median ocellus
becomes illuminated directly at point (a) subsequently, in the early noon hours
(and in the course of its orientation flight), direct illumination of its median
ocellus shifts to point (b) and later on to point (c), associated with the late
noon hours and finally, toward evening and twilight -to point (d). Note that
all the afore said pertains also to the two lateral ocelli as well. When the
hornet makes its rotation clockwise the situation is reversed, as observable
in Fig. 5.
Maturation and first flight movements of the hornet
When a hornet eclodes, it is rather pallid and even its future brown and yellow
zones are still pale. This means that the vespan cuticle and perhaps also its
sense organ and other organs undergo 'ripening' during full maturation of the
insect. Neither is the fledgling hornet properly orientated to its environment,
so that upon its removal to some distance from the nest, it is incapable of
finding its way back. Undoubtedly, for a hornet to fulfill its various tasks,
it needs to reach 'maturity' in certain functions following its eclosion. Such
maturation transpires probably within a few days. In my experience, it is a
matter of about five days, during which there is: (1) tanning of the cuticle;
(2) maturation of the various sense organs, lending the insect proper orientation
to its environment and ensuring that once it leaves the nest it will know how
to find its way back. My preliminary observations indicate that during the first
few days post eclosion, the young hornets venture outside the nest to short
distances of several meters only. Their flight during these few days is slow
and unsteady, mostly in circles around the nest. It is only gradually that the
hornets venture to fly greater distances. I presume that during this interim
period the hornet learns to orientate the nest location with the varying direction
of the sun in the sky during the changing hours of the day. This learning process
is a sine qua non for ultimate orientation of hornets during foraging flights.
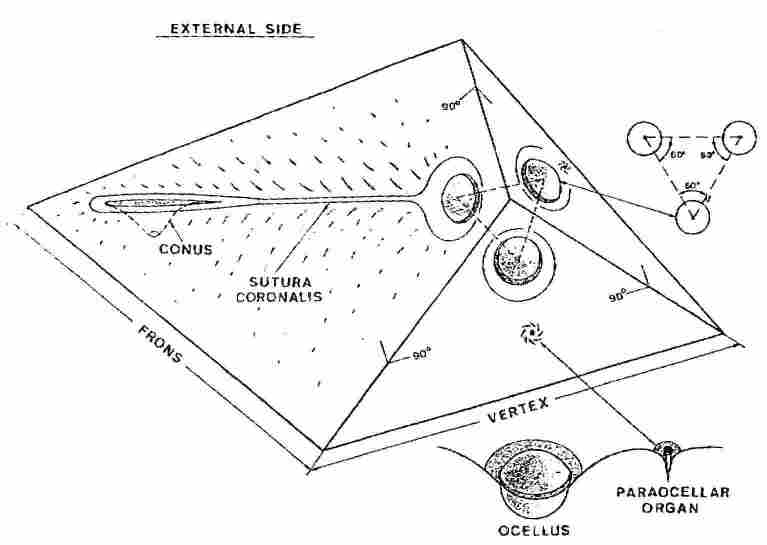
Fig. 1. Scheme showing the location of the ocelli, conus and
paraocellar organs on the vertex and frons. Each ocellus is situated on a separate
plate, with the angle between the plates being 90° . The three plates thus form
a triangular pyramid whose front facet bears the median ocellus and its continuation
bears also the frons plate with the coronal suture in its center and behind
it the conus and the "Ishay Organ". At the back of each lateral ocellus there
is a paraocellar organ. At the bottom of Fig. 1 can be seen an ocellus with
a paraocellar organ behind it. Note that the ocellus is spherical and surrounded
by a canal. If one draws an imaginary triangle between the three ocelli, this
will be an equilateral triangle with an angle of 60° at each corner.
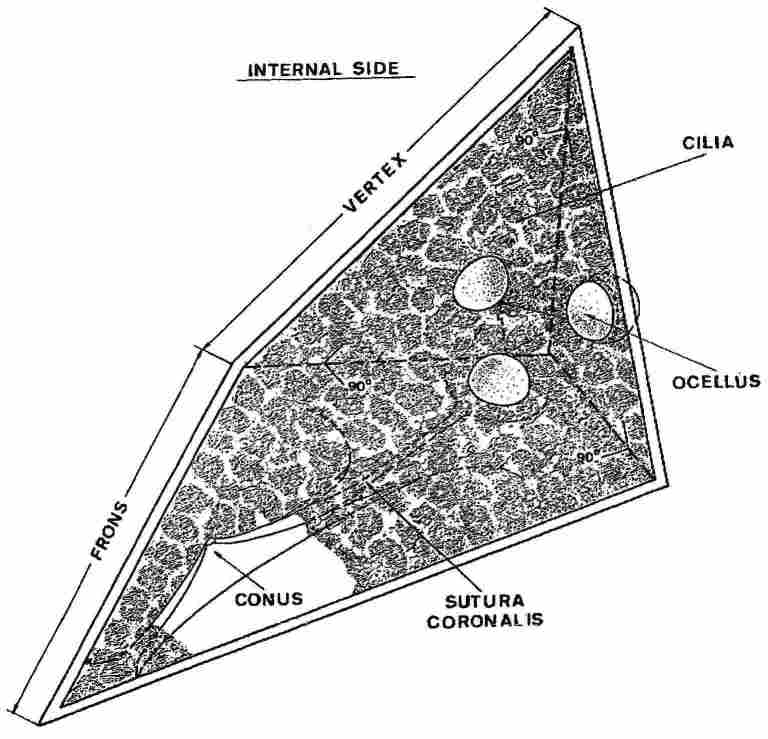
Fig. 2. View of the inner sides of the frons and vertex plates. On right, one
sees the three ocelli and on the left the conus. The entire surface, barring
the ocelli, is covered with hair cells from which protrude the cilia.
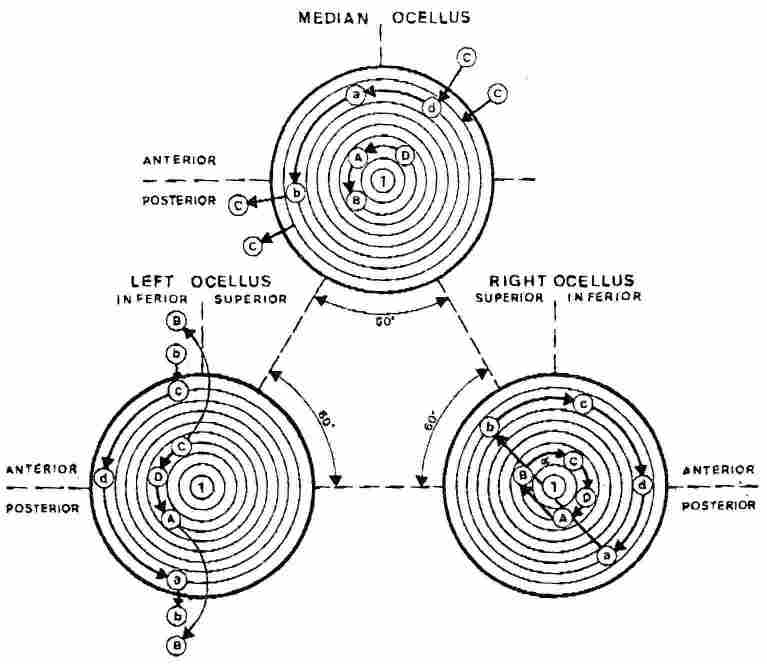
Fig. 3. Scheme of the changing trajectory of sunlight on the
vespan ocelli during daytime flight. The indicated trajectory corresponds to
a counterclockwise rotary movement of the hornet. Capital letters designate
the trajectory during the early morning and late evening hours, whereas small
letters designate the pattern during the noon hours.
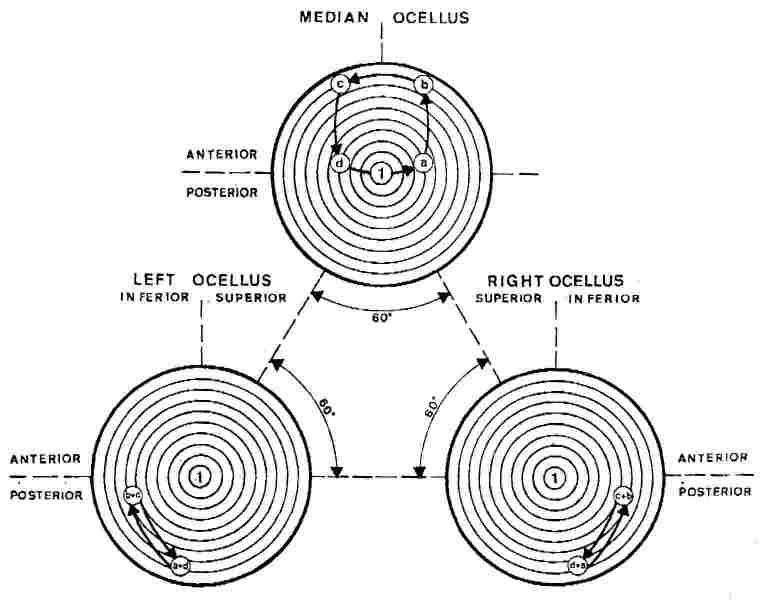
Fig. 4. Scheme depicting the path of the sun rays on the ocelli
during daytime hours, with the head of the flying hornet orientated towards
the sun. In this situation, the median ocellus is illuminated in symmetrical
fashion. As can be seen , the trajectory of the sun rays is from the center
in the morning hours to the periphery in the noon hours and back to the center
in the late afternoon hours

Fig. 5. Schematic representation of the vertex with the three
ocelli, as it relates to sun direction and the zenith. 1 = sun and 2 = zenith.